
Ein Post auf Uncommon Descent (ausgerechnet) hat mich an ein Thema auf meiner "Liste von Dingen, über die ich unbedingt was schreiben will (irgendwann, bestimmt...)" erinnert: Die molekularen Mechanismen der Entstehung neuer Gene und Genfunktionen.
Dazu gibt es eine hervorragende Zusammenfassung in Nature Reviews Genetics [1]. Die verschiedenen darin erwähnten Mechanismen der Entstehung neuer Genfunktionen sind die Folgenden:
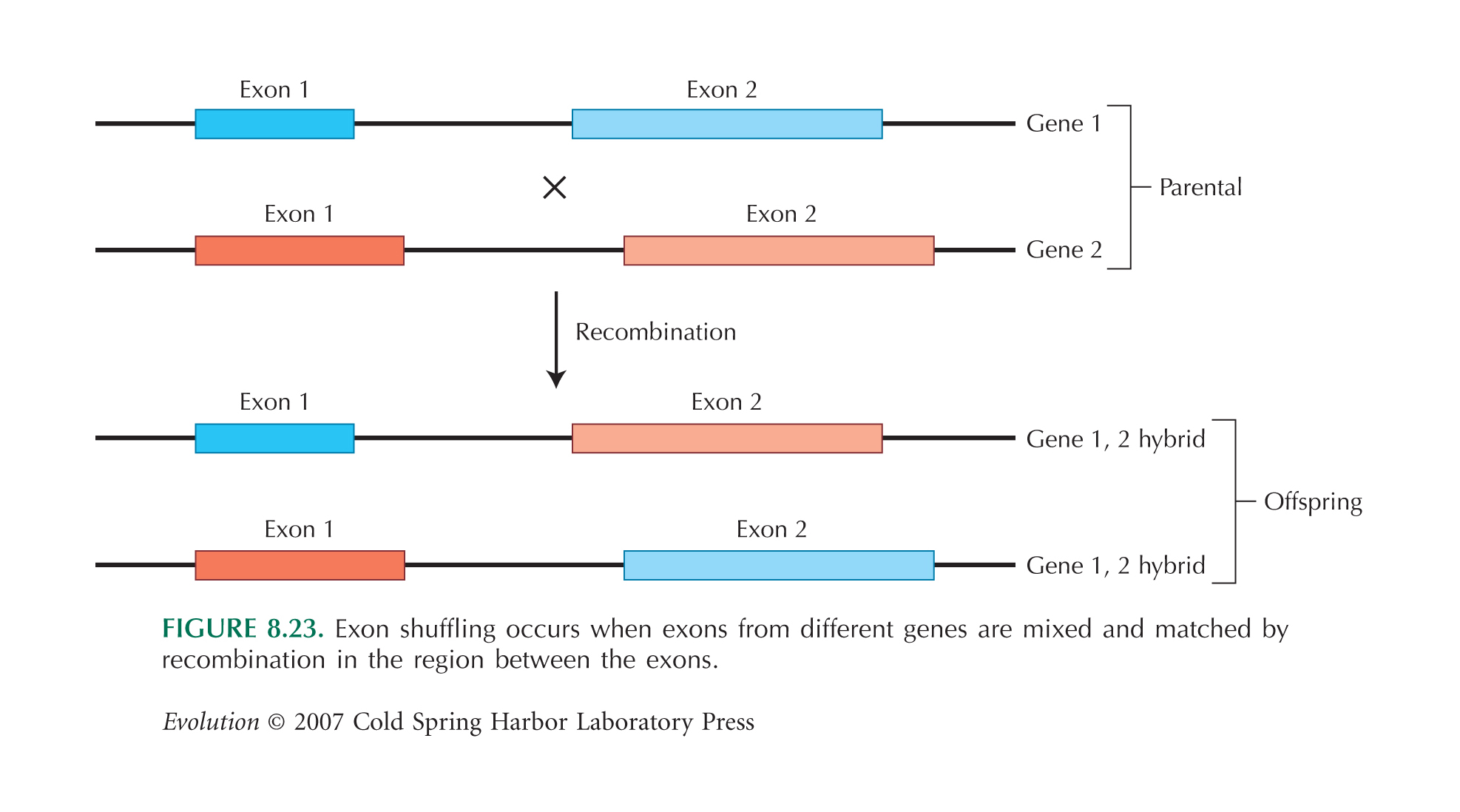
Exon shuffling: Bei Eukaryoten sind die proteinkodierenden Abschnitte der Gene (Exons) von nicht-kodierenden Sequenzen, den Introns, unterbrochen. Exon shuffling ist die Neukombination von Exons verschiedener Herkunft, also aus verschiedenen Genen. Dies kann z. B. durch Crossing over, "ungenaue" Rekombination (d. h. es findet ein Austausch zwischen nicht homologen Bereichen des Genoms statt [siehe Abbildung hier; aus [2]) oder durch Retrotransposition (s. u.) geschehen.
Es wird angenommen, dass etwa 19 % aller Exons in eukaryotischen Genen schon einmal ein Exon shuffling "mitgemacht" haben.
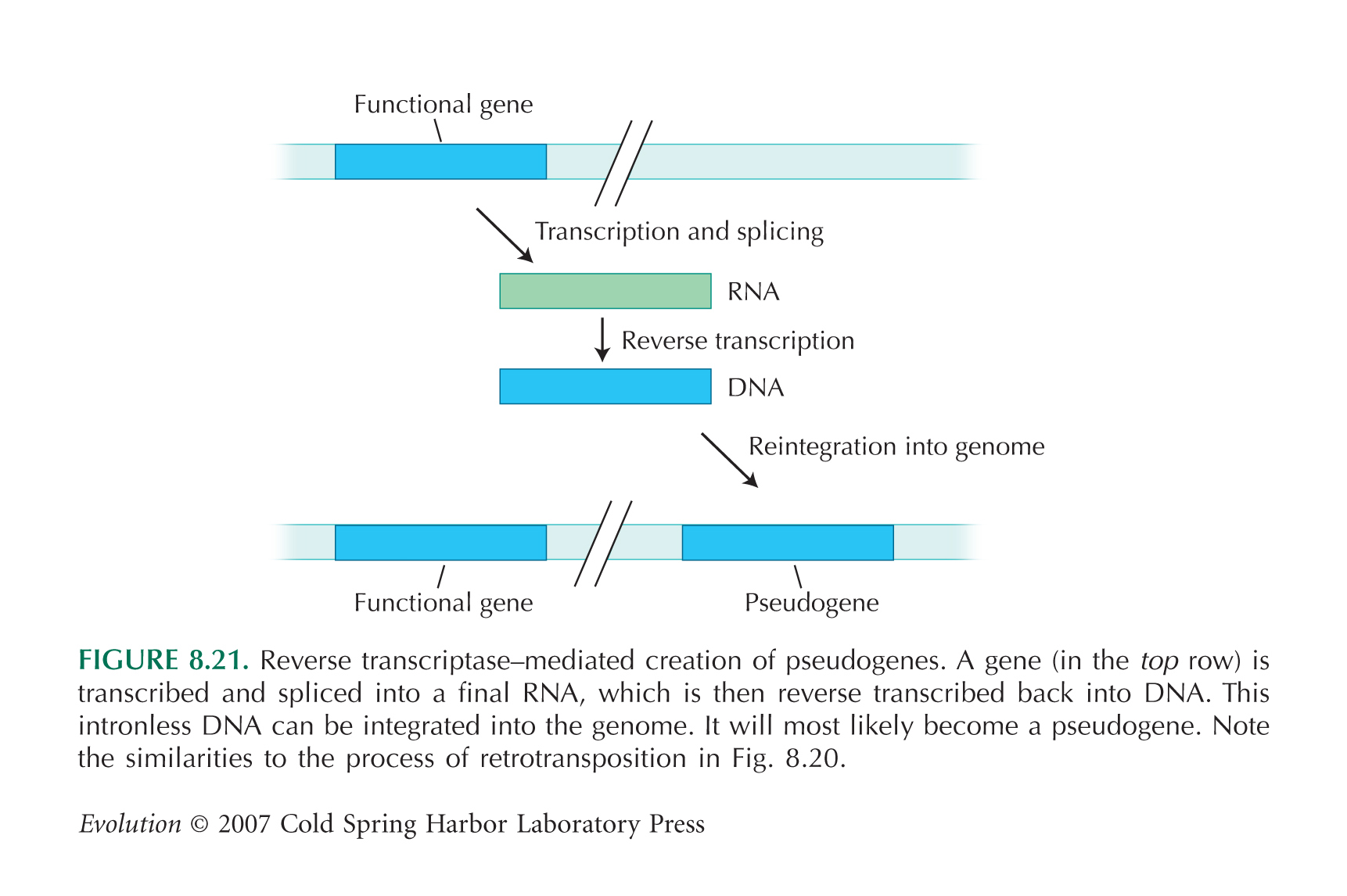
Gen-Duplikation: Der wohl bekannteste Mechanismus, durch den neue Gene entstehen können, ist die Duplikation vorhandener Gene, mit einer anschließenden Diversifizierung einer der beiden Genkopien, wodurch neue Funktionen entstehen können. Ursache können wieder z. B. Retrotransposition (s. u.), nicht-homologe Rekombination oder Ablesefehler sein. Bei der Retrotranposition wird nur der Protein-kodierende Teil verdoppelt. Damit das "doppelte" Gen abgelesen werden kann, müssen an der Integrationsstelle genregulierende Elemente vorhanden sein. Schon allein durch eine von dem Ursprungsgen verschiedene Genregulierung können neue Genfunktionen entstehen.
Aber auch ganze Chromosomen-Abschnitte oder ganze Genome können verdoppelt werden. Bei Pflanzen ist die Verdoppelung ganzer Genome relativ häufig, bei den Angiospermen (den "blühenden" Pflanzen) entstehen etwa 2-4 % aller Arten auf diese Weise. Da sich Tiere in der Regel sexuell fortpflanzen und daher mindestens zwei Individuen mit verdoppeltem Genom vorhanden sein (und aufeinander treffen) müssen, um eine neue Art zu gründen, ist dies im Tierreich nicht so häufig, ist aber auch schon mehrfach vorgekommen [2].
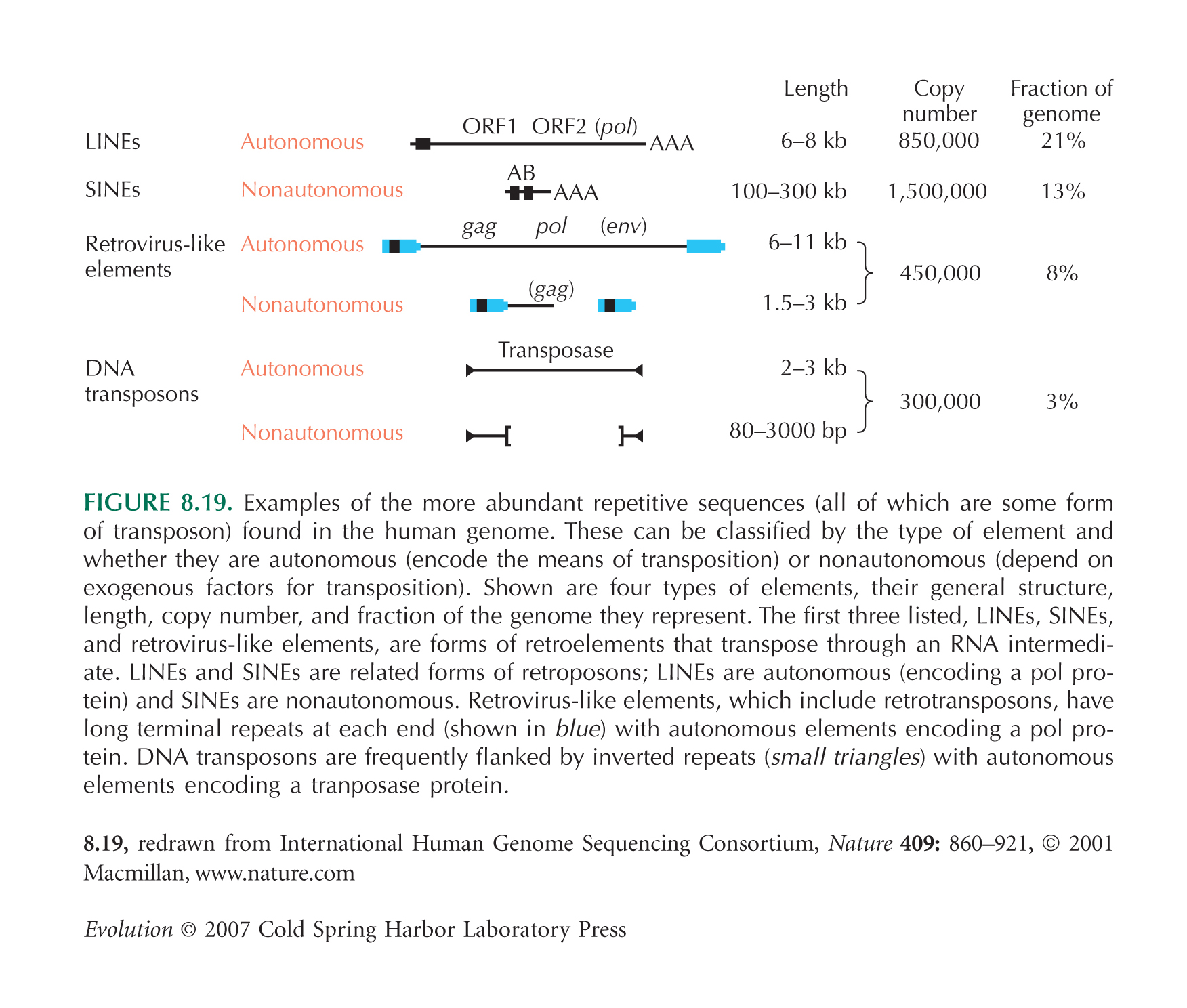
Retrotransposition: Retrotransposons sind genomische Elemente (beim Menschen beispielsweise das sog. LINE-1- oder L1-Element; für die Häufigkeiten verschiedener Transposons siehe z. B. hier), die mindestens zwei Gene kodieren, eine Endonuklease (ein Enzym, dass doppelsträngige DNA schneiden kann) und eine Reverse Transkriptase (ein Enzym, das RNA in DNA umschreiben kann). Retrotransposons "vermehren" sich, in dem sie zunächst durch RNA-Polymerase II in RNA umgeschrieben werden. Die enthaltenden Protein-kodierenden Sequenzen werden ganz normal an Ribosomen in Aminosäuren "übersetzt", so dass die oben genannten Enzyme bereitstehen. Diese verbinden sich mit der L1-RNA, schreiben sie in DNA um und integrieren diese L1-DNA-Sequenz irgendwo im Genom. "Versehentlich" kann auch andere RNA, also auch die mRNA anderer exprimierter Gene (oder Teile davon, beispielsweise auch nur ein Exon) in DNA umgeschrieben und ins Genom integriert werden (für eine vereinfachte Abbildung siehe hier; aus [2]). Retrotransposition ist einer der Hauptursachen für Genduplikationen (die duplizierten Gene sind daran zu erkennen, dass sie keine Introns enthalten), aber auch für Exon shuffling, wenn nur ein Teil eines Gens in DNA umgeschrieben wurde und in ein anderes Gen integriert wurde.
Durch das L1-Element verursachte Retropositionen waren an der Entstehung von etwa 1 % des menschlichen Genoms beteiligt.
Mobile Elemente/Transposons: Transposons sind ebenfalls genetische Elemente ähnlich den Retrotransposons. Anders als diese kodieren sie keine Reverse Transkriptase. Transposons können sich an einer Stelle des Genoms ausschneiden und an anderer Stelle wieder integrieren. Wenn sie sich innerhalb eines Gens integrieren, können sie neue Exons bilden. Etwa 4 % aller neuen Exons im menschlichen Genom gehen auf Transposons zurück.
Lateraler/horizontaler Gentransfer: Gene werden von einen Organismus auf den anderen übertragen, beispielsweise durch Viren, Bakteriophagen, durch Plasmide oder durch Endoparasiten. Lateraler Gentransfer kommt besonders häufig bei Prokaryoten vor, ist aber auch schon bei Eukaryoten beobachtet worden. Beispielsweise wurde kürzlich entdeckt, dass der Endosymbiont Wolbachia fast sein gesamtes Genom auf seinen Wirt, Drosophila ananassae, übertragen hat [3].
Genfusion oder Genaufspaltung: Neue Gene können entstehen, wenn aus zwei Genen eins wird (beispielsweise, in dem das Stopcodon des "ersten" Gens durch Mutation zerstört wird und bis in das zweite "durchgelesen" wird. Seltener nachgewiesen ist die Aufspaltung eines Gens, d. h. es entstehen zwei kürzerer Gene.
De novo-Entstehung: Gene können aus nicht-kodierenden Sequenzen entstehen.
Alle diese Mechanismen der Genentstehung sind nachgewiesen und durch viele Beispiele belegt. Die neu entstandenen Strukturen unterliegen teilweise einer rasanten Evolution. Nach Genduplikationen wurde z. B. bei Drosophila eine mindestens zehnfach schnellere Veränderung der "neuen" Gensequenz verglichen mit der "Original"-Gensequenz festgestellt, Gleiches wurde auch im menschlichen Genom beobachtet [1].
Bisher ist man davon ausgegangen, dass eine "echte" De novo-Entstehung von Genen aus vorher nicht-kodierenden Sequenzen nur sehr selten vorkommt. Dies wurde aber nicht wirklich systematisch untersucht. Jetzt ist ein Artikel in Genome Research [4] erschienen, der sich des Themas angenommen und in verschiedenen Drosophila-Arten nach neu entstandenen Genfunktionen und Genen gesucht hat.
Das Abstract fast die Ergebnisse sehr schön zusammen:
Several mechanisms have been proposed to account for the origination of new genes. Despite extensive case studies, the general principles governing this fundamental process are still unclear at the whole genome level. Here we unveil genome-wide patterns for the mutational mechanisms leading to new genes, and their subsequent lineage-specific evolution at different time nodes in the D. melanogaster species subgroup. We find that, 1) tandem gene duplication has generated about 80% of the nascent duplicates that are limited to single species (D. melanogaster or D. yakuba); 2) the most abundant new genes shared by multiple species (44.1%) are dispersed duplicates, and are more likely to be retained and be functional; 3) de novo gene origination from non-coding sequences plays an unexpectedly important role during the origin of new genes, and is responsible for 11.9% of the new genes; 4) retroposition is also an important mechanism, and had generated approximately 10% new genes; 5) about 30% of the new genes in the D. melanogaster species complex recruited various genomic sequences and formed chimeric gene structures [Anm.: chimeric gene structures können beispielsweise aus Fusionen von kodierenden Regionen und Intronsequenzen zweier seperater Gene entstehen, eine Mischung aus Exon shuffling und Genfusion], suggesting structure innovation as an important way to help fixation of new genes; and 6) the rate of the origin of new functional genes is estimated to be 5 to 11 genes per million years in the D. melanogaster subgroup. Finally, we survey gene frequencies among 19 strains from all over the world for D. melanogaster-specific new genes, and reveal that 44.4% of them show copy number polymorphisms within population. In conclusion, we provide a panoramic picture for origin of new genes in Drosophila species.Wirklich überraschen ist der hohe Anteil an neuen Genen, die wirklich de novo entstanden sind: fast 12 %! Wo das doch laut IDlern völlig unmöglich ist...
MfG,
JLT
[1] Long et al. (2003). The Origin of new genes: glimpses from the young and old. Nature Review Genetics 4: 865-875.
[2] Barton et al. (2007). Evolution. Cold Spring Harbor Laboratory Press.
[3] Hotopp et al. (2007). Widespread lateral gene transfer from intracellular bacteria to multicellular eukaryotes. Science 317: 1753-1756
[4] Zhou et al. (2008). On the origin of new genes in Drosophila. Genome Research, published online Jun 11, 2008 [doi:10.1101/gr.076588.108]













{kind=link}
{kind=link}
{kind=link}
1 Kommentare:
Hi,
Ich habe neulich über das neuentdeckte Mamavirus und das erste Virus-Virus, genannt Sputnik, geposted. Dessen gene sind zu grossen Teilen einfach vom Wirt "geklaut". Auch eine brauchbare Methode!
Grüsse
Post a Comment